 Schulz begynner med å liste opp de forskjellige delene av flagellen, og beskriver deres funksjonelle-mekaniske-biokjemiske forhold. Schulz begynner med å liste opp de forskjellige delene av flagellen, og beskriver deres funksjonelle-mekaniske-biokjemiske forhold.
Schulz begynner med å liste opp de forskjellige delene av flagellen, og beskriver deres funksjonelle-mekaniske-biokjemiske forhold. Schulz begynner med å liste opp de forskjellige delene av flagellen, og beskriver deres funksjonelle-mekaniske-biokjemiske forhold. 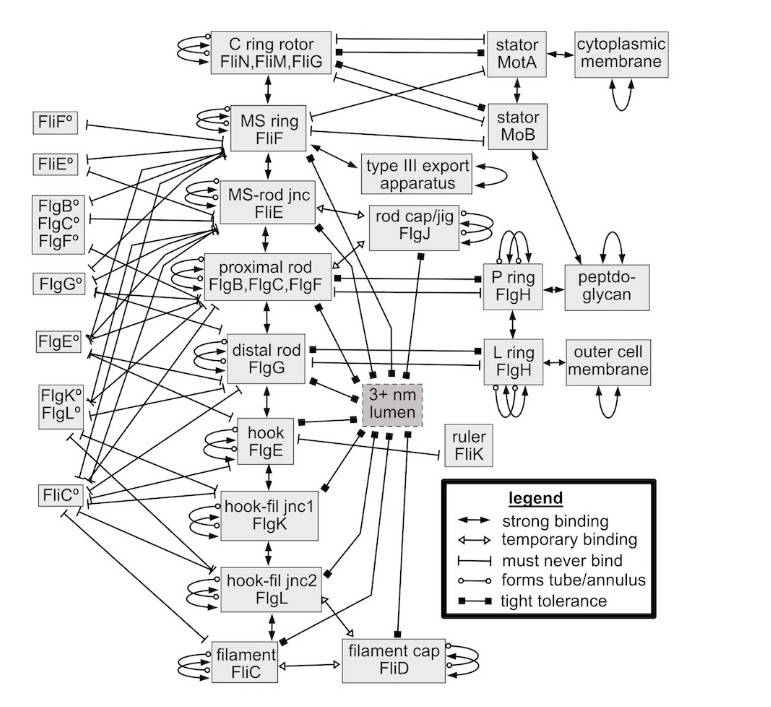 Linjene i figur 1 representerer svært spesifikke egenskaper til proteinene som utgjør de forskjellige underenhetene. For det første må den molekylære strukturen til proteinene som danner en ringrom eller rør, ha egenskapen at proteinene har bindingssteder som forbinder proteinene godt slik at de sekvensielt støter mot hverandre. Ved å gjøre det må hvert protein effektivt danne en N-graders bue. Det er en veldig spesifikk geometrisk og aminosyresekvens-egenskap for et foldet protein, som bare ekstremt sjeldne proteinkonfigurasjoner kan møte. Videre, for proteiner som danner et rør, binder hver runde av de spiralformede, kveilede-tau-lignende, ende-til-ende-koblede proteinene, og hver løkke av helixen til de foregående og etterfølgende løkkene for å danne et stabilt rør, og kan dermed overføre dreiemoment effektivt ved å være tangentielt stiv. Med andre ord må mange av proteinene binde seg til fire andre tilstøtende proteiner: proteinene foran og bak langs hver løkke av helixen, og proteinene i de foregående proksimale og påfølgende løkkene rundt helixen. De nærliggende proteinene kan være like (som i filamentet) eller forskjellige (hvor en underenhet binder seg til den neste). Det vil si at proteinene til hver delenhet må binde seg fast til proteinene fra minst én annen delenhet. Disse bindingsegenskapene krever strenge, veldig spesifikke krav til den foldede kjeden av aminosyrerester til disse proteinene, det vil si de innrettede stedene for ikke-kovalent binding mellom to like eller forskjellige proteiner.
Linjene i figur 1 representerer svært spesifikke egenskaper til proteinene som utgjør de forskjellige underenhetene. For det første må den molekylære strukturen til proteinene som danner en ringrom eller rør, ha egenskapen at proteinene har bindingssteder som forbinder proteinene godt slik at de sekvensielt støter mot hverandre. Ved å gjøre det må hvert protein effektivt danne en N-graders bue. Det er en veldig spesifikk geometrisk og aminosyresekvens-egenskap for et foldet protein, som bare ekstremt sjeldne proteinkonfigurasjoner kan møte. Videre, for proteiner som danner et rør, binder hver runde av de spiralformede, kveilede-tau-lignende, ende-til-ende-koblede proteinene, og hver løkke av helixen til de foregående og etterfølgende løkkene for å danne et stabilt rør, og kan dermed overføre dreiemoment effektivt ved å være tangentielt stiv. Med andre ord må mange av proteinene binde seg til fire andre tilstøtende proteiner: proteinene foran og bak langs hver løkke av helixen, og proteinene i de foregående proksimale og påfølgende løkkene rundt helixen. De nærliggende proteinene kan være like (som i filamentet) eller forskjellige (hvor en underenhet binder seg til den neste). Det vil si at proteinene til hver delenhet må binde seg fast til proteinene fra minst én annen delenhet. Disse bindingsegenskapene krever strenge, veldig spesifikke krav til den foldede kjeden av aminosyrerester til disse proteinene, det vil si de innrettede stedene for ikke-kovalent binding mellom to like eller forskjellige proteiner.
FRA TOPPEN OG NED -spesifikasjon |
FRA BUNNEN OG OPP -analyse |
nødvendige fremdriftsfunksjoner |
arketypisk implementering |
kraftkilde |
ion (H+ eller natrium) gradient |
kraft-til-bevegelse virkemiddel |
roterende elektrisk motor |
ytre komponent(er) |
stang-ende, krok, filament |
fundament/substrat |
indre/ytre membraner, peptidoglykan |
roterende delsystem |
se diagram (over) |
anker |
MS-ring: FliF-proteiner |
motorrotor |
C-ring: FliG, FliM, FliN |
Akselstang: |
aksel: FlgB, FlgC, FlgF; FlgG |
aksial bøying, hvis sidemontering |
krok: FlgE |
spiralformet propellfilament: |
FliC, FliD (hette) |
(mulige adaptere) |
krok-filament: FliE, FlgK, FlgL, FlgM |
tetningslager |
P-ring & L-ring: FlgI, FlgH |
motorstator |
MotA, MotB |
 Diverse perspektiver på flagellen
Diverse perspektiver på flagellen Molekylærbiologen er mindre opptatt av opprinnelse enn av hvordan systemer fungerer.
Molekylærbiologen er mindre opptatt av opprinnelse enn av hvordan systemer fungerer. 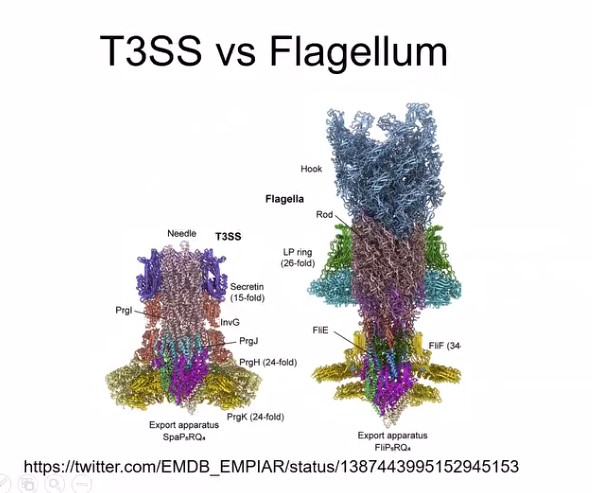 Evolusjonære hindringer og intelligent design
Evolusjonære hindringer og intelligent design Schulz er i tvil om at blind engineering er mulig - men han later ikke som om alle spørsmål er besvart:
Schulz er i tvil om at blind engineering er mulig - men han later ikke som om alle spørsmål er besvart:
Oversettelse og bilder ved Asbjørn E. Lund